|
|
|
| Click here to visit our web site |
|
INTRODUCTION CHONDROGENESIS DURING EMBRYONIC DEVELOPMENT
Decades of work in chick, mouse, and other models have delineated several functional classes of genes that mediate aspects of embryonic development. A “master gene” paradigm has emerged for embryonic cell differentiation, in which induction of a specific DNA binding protein alters gene expression to bring about the differentiated phenotype. The prototype system is the mechanism of action of the basic helix-loophelix protein, myoD, during differentiation of muscle cells. The “master gene” paradigm is emerging for other tissues of mesenchymal origin: the runt-domain protein Osf2/Cbfa-1 is essential for osteoblast development, and the nuclear hormone receptor PPAR γ is required for development of adipose cells. Accordingly, there has been a search for a chondrogenic “master gene”. There are several candidate genes, including the HMG-box protein, Sox9; the bHLH protein, scleraxis; and the homeobox protein, Hoxc8. Peptide growth factors are another functional class of genes that influence embryonic chondrogenesis. TGF-α and its family members GDF-5 and the BMPs are essential for prenatal chondrogenesis. Several members of the Wnt family of secreted glycoproteins have been shown affect embryonic chondrogenesis. The extracellular matrix can also influence chondrocyte differentiation and chondrogenesis during embryonic development. Null mutations in the mouse col2a1 or colXa1 genes result in abnormal cartilage elements. Mice carrying a null mutation for perlecan, a heparin sulfate proteoglycan, have defective growth plates and disorganized cartilage matrix. A comparison of chondrogenic mechanisms in avian and mammalian development shows that there are some speciesspecific differences. For example, a loss-of-function mutation in the mouse wnt5a gene generates pronounced skeletal defects: there is shortening of the limb, and digits fail to form. These observations support an instructive role for Wnt5a in specifying chondroblast differentiation in mammalian limb development. In the embryonic chick limb, however, overexpression of Wnt5a retards growth by delaying or inhibiting maturation of prehypertrophic chondrocytes. This suggests that, in the chick, Wnt5a is a suppresser of chondrogenesis. How do the mechanisms of prenatal chondrogenesis compare to those of postnatal chondrogenesis? It has been difficult to address this question experimentally because, in a normal individual, postnatal chondrogenesis does not occur outside of the growth plate or the soft callus of fracture repair. If postnatal chondrogenesis is affected by genes other than those associated with prenatal chondrogenesis, opportunities for therapeutic intervention may be greater than originally thought. IN VITRO MODEL OF POSTNATAL CHONDROINDUCTION
IDENTIFICATION OF DIFFERENTIALLY EXPRESSED
GENES IN CHONDROINDUCED FIBROBLASTS
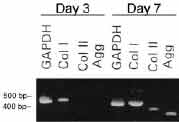
As a first step toward identifying mechanisms of postnatal chondroblast differentiation, we analyzed shifts in gene expression that occur in hDFs cultured in DBP/collagen sponges. To determine an appropriate timepoint for gene expression analysis, we evaluated evidence of chondroblast differentiation in DBP/collagen sponges cultured for 3 and 7 days. After 3 days, hDFs were distributed throughout the sponges and were attached to the collagen lattice; those cells that had migrated into the packet of DBP were attached to, and between the particles of DBP. Metachromatic extracellular matrix was not observed in the DBP/collagen sponges at 3 days. After 7 days, however, metachromatic extracellular matrix and cartilage proteoglycans were present. (9) Moreover, expression of cartilage-specific genes was not detectable in hDFs cultured in DBP/collagen sponges for 3 days, but was induced by 7 days (Figure 1). Three days was therefore taken to represent a timepoint at which early interactions were occurring between the cells and DBP, prior to expression of the chondroblast phenotype. We used representational difference analysis, a PCR-based method of subtractive hybridization, to isolate a pool of genes whose expression was increased (Upregulated) in chondroinduced hDFs three days after exposure to DBP. 9 Several of the genes we identified were novel. Others corresponded to mRNA transcripts of unknown cellular function. Many Upregulated genes, however, corresponded to genes with known functions, including protein synthesis, cytoskeletal elements, extracellular matrix, and the regulation of gene expression. Several aspects of each cellular process were represented among the functional classes of Upregulated genes. For example, the synthesis, aminoacylation, and export of tRNA from the nucleus are all required steps in protein synthesis. Different types of cytoskeletal elements were also identified: gene products associated with the actin- or microtubule-based cytoskeleton, gene products involved in cell adhesion, and gene products that function in messenger RNA attachment to the cytoskeleton. Genes involved in the regulation of gene expression fell into two classes: transcription factors and DNA helicases. Transcription factors provide gene-specific control of expression. In contrast, DNA helicases unwind strands of chromatin, thereby enabling access of transcription factors to their DNA sequence-specific binding sites. Helicases can also prevent transcription by re-winding chromatin. SHIFTS IN GENE EXPRESSION IN CHONDROINDUCED
FIBROBLASTS The observed gene expression patterns, together with histologic and biochemical evidence of the chondroblast phenotype, suggest that there are specific stages of induced chondrocyte differentiation in our experimental system (Figure 2). Three days after exposure to DBP, the cell is between induction and overt differentiation. Gene expression shifts are already detectable in this early, determined neochondroblast stage. Elevated expression of some genes is transient. Elevated expression of other genes continues while cartilage-specific matrix is being actively synthesized in the differentiated neochondroblast stage. Cartilage-specific matrix gene expression, which is present at the differentiated neochondroblast stage, continues to increase until the cell reaches the mature stage of a chondrocyte.
CONCLUSIONS
ACKNOWLEDGEMENTS Karen E. Yates, PhD, Instructor, Harvard Medical School, Orthopedic Research Laboratory, Brigham and Women’s Hospital. Address Correspondence to: |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Print Manuscript • View References • Download PDF version • Close window |