| HOJ HOME | Chiefs Reports | Osgood Day | Scientific Articles | Alumni Association |
|
| Click here to visit our web site |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Cooperative interactions
between bone forming osteoblasts and bone-resorbing osteoclasts have been
a central focus of orthopaedic research for almost two decades, following
the landmark hypothesis advanced by G. Rodan and T. J. Martin in 1981
(1) . A perplexing problem and major impediment to studying osteoclasts
was the experimental observation that physical contact between mononuclear
osteoclast precursor cells and stromal "fibroblasts" was required for
the differentiation of functional multinucleated osteoclasts in bone marrow
cultures. The solution to this problem, reviewed below, represents an
exciting breakthrough with important applications to orthopaedic research
and clinical practice.
The
New Pathway for Osteoclast Formation: Discovery of the RANK / RANKL Complex
Osteoclast differentiation
and activation was recently proven to be dependent on formation of the
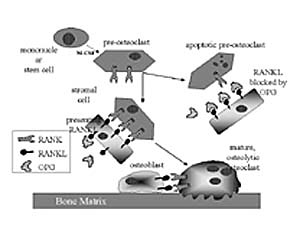
RANK/RANKL receptor/ligand complex (Table 1, Fig. 1) (2) . RANK (Receptor
Activator of NF-KappaB) was discovered in 1997 in dendritic cells (antigen-presenting
cells of the immune system), and was shown to be critical for their survival
during interaction with T-helper cells (3) . The novel ligand that binds
to RANK was cloned and named RANKL ("RANK Ligand") by the Immunex research
group (3) and TRANCE by another lab (4) . Independently, osteoclast research
teams lead by T. Suda and H. Yasuda cloned osteoclast differentiation
factor (ODF) and its receptor (ODFR), which proved to be identical to
RANKL and RANK, respective-ly (Table 1) (2) . RANK is a type II membrane
protein in the tumor necrosis factor receptor (TNFR) family, and is related
to CD40, Fas, TNFR1 (p55), TNFR2 (p75), OX40, and to OPG/OCIF, the novel
bone-protecting decoy receptor for RANKL discussed below. The TNFR family
operates on a highly specific one receptor-one ligand principle, and RANKL
apparently serves as the obligate ligand for RANK (3) . RANKL is homologous
to other TNFR family ligands (TNF , FasL, CD40L, TRAIL), some trig-gering
and others protecting against apoptosis, depending on the recruitment
of death domain proteins and TNFR-associat-ed factors (TRAF) in the target
cell.
Rationale for Studying the RANK / RANKL / OPG System in Skeletal Disease This pathway for osteoclast development provides a con-vergent, unified mechanism for understanding the regulatory actions of many disparate agents (1,25(OH)2 -vitaminD (3) , PTH/PTHrp, IL-6, IL-11, M-CSF, and OPG) in skeletal homeostasis, and pathology (e.g., metabolic bone disease, inflammatory bone destruction, and osteolytic metastasis). Novel interpretations of earlier studies on skeletal metastasis are now possible in light of the hypothesis that we and others are testing: namely that the RANK / RANKL / OPG pathway is utilized by breast adenocarcinoma to establish osteolytic sites. A recent study suggests that breast cancer-derived PTHrp, acting on osteoblasts to upregulate RANKL expression, is a major stimulus to osteoclast formation in situ (9) . Future therapies for bone metastases will undoubtedly target this new class of tumor-bone interactions and the regulatory mechanisms involving RANK, RANKL, and OPG. Model Systems for Osteoclast Study With the availability of recombinant soluble forms of RANKL that can act on RANK without being tethered to a membrane (3) , osteoclasts can now be formed efficiently in vitro. Our attention has recently focused on the mouse macrophage cell line RAW264.7 that expresses RANK and responds strongly to mRANKL, forming TRAP+ multinucleated osteoclasts within 4 days (7) . A newly developed resorbable hydroxyapatite film culture system allows us to study resorptive activity of individual osteoclasts. Importantly, the RAW osteoclasts can be transfected in pure culture for gene promoter studies and the possible discovery of novel osteoclast-specific transcription factors (see below). Transcriptional Regulation in Osteoclasts Maturation of osteoclast precursors involves the expression of several osteoclast enriched and osteoclast specific proteins: structural proteins, receptors (e.g. calcitonin receptor, RANK), signaling molecules (e.g. Src), enzymes (e.g. cathep-sin- K, tartrate-resistant acid phosphatase, osteoclast proton-pump components, and carbonic anhydrase), membrane channels (e.g. Cl - channel and Cl - /HCO 3 - anion exchanger), and adhesion molecules (e.g. á V â 3 integrin) (10) . The integrin á V â 3 is a member of a large family of heterodimeric, trans-membrane, cell-matrix attachment proteins which recognize specific sequences in a variety of extracellular matrix proteins. Integrins not only mediate cell-matrix attachment but also transmit bi-directional signals across the plasma membrane (11) . Integrin adhesion and signal transduction con-tribute to a wide range of cellular activities including motility, spreading, differentiation, secretion of matrix proteins, matrix assembly, matrix mediated cell cycle progression, and apoptosis (11) . Mature osteoclasts predominantly express the á V â 3 form of integrin, with levels estimated at 10 to the 7th molecules per osteoclast (12) . Pharmacological blockade of ligand binding, by the highly expressed á V â 3 osteoclast integrin, inhibits osteoclast adhesion to bone and to specific bone matrix proteins, while inhibiting the spreading of osteoclasts. Osteoclast functional activity, as measured in vitro by pit formation and resorption of radioactive bone fragments, is also inhibited by blockade of á V â 3 binding (13) . In addition, ovariectomy-induced bone loss is inhibited by blockade of á V â 3 in animal models of post-menopausal osteoporosis (14, 15) . The requirement for â 3 integrin expression in osteoclast formation and function in vivo was determined by investigating mice in whom the â 3 gene was genetically deleted. Mice that lack the â 3 gene, and therefore do not express the á V â 3 integrin, produce defective osteoclasts. Osteoclasts from â 3 null mice do not spread normally in vitro, do not form the characteristic actin ring structure, and have disorganized ruffled membranes in vivo. These defects, while countered by a 3-fold increase in osteoclast numbers, lead to increased bone mass and osteosclerosis, confirming the importance of á V â 3 for normal osteoclast function (16) . Expression of the á V â 3 integrin heterodimer is regulated at the level of the â 3 integrin mRNA, with no significant changes in á V mRNA levels during hormone and cytokine induced á V â 3 expression in macrophages and osteoclast precursors (17-19) . The increase in â 3 integrin mRNA steady state level occurs through transcriptional up-regulation and not via increased message half-life. Importantly, the huge increase of á V â 3 expression during osteoclast formation results from a transcriptional increase in â 3 integrin mRNA levels (19) . Little is known about the regulation of gene transcription during osteoclastogenesis, primarily because a tractable cell model system for osteoclast formation has not been available. Recent advances in our understanding of the signals and factors directing osteoclastogenesis led to the identification of such a model cell line, RAW264.7 (7 ). To study the regulation of â 3 gene expression in osteoclastogenesis, we generated â 3 gene promoter/luciferase (luc) gene reporter constructs. In tran-sient transfection with these constructs, expression of the light producing luc gene reporter is a direct measure of the â 3 gene promoter activity. By transient transfection of osteoclasto-genic cells with the â 3 promoter/luc reporter construct, and subsequent induction of osteoclast formation with RANKL, we have shown that a 1.1 kilobase fragment of the â 3 promoter directs an 7 to 11 fold increase in reporter expression [McHugh, unpublished]. Deletion analysis of the 1.1 kilobase promoter have further identified an approximately 65 base pair region containing sequences which are responsible for the bulk of promoter induction during osteoclast formation. Further deletion and mutation of the â 3 promoter, in luc reporter con-structs, will allow us to identify the DNA sequences and their cognate transcription factors that direct â 3 gene transcription in osteoclastogenesis.
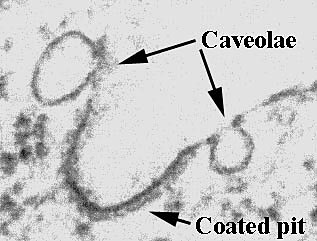
Caveolae and Pre-assembled Signal Transduction Complexes in Osteoblasts Signal transduction generally begins with the binding of a ligand to a membrane-anchored receptor, initiating a cascade of intracellular events leading to protein phosphorylation, protein- protein interaction, gene regulation, and a variety of cellular changes or responses. Signaling events in pre-osteoclasts that lead to osteoclast maturation (see above) often originate from osteoblasts that are actively responding to hormones or cytokines with their own specific set of signal transduction pathways. Thus it is essential to understand the signaling events in osteoblasts. Recently, we demonstrated that osteoblasts have specialized pre-assembled signal transduction complexes and caveolae (Fig. 2), tiny membrane structures enriched with signaling molecules. Before describing the form and function of caveolae and pre-assembled signal transduction complexes in osteoblasts, we will address the origin of the membranes and nature of the lipids and proteins that form caveolae. Small Detergent-resistant Membrane Domains (Detergent-resistant Microdomains) Sphingolipids and cholesterol form detergent-resistant membrane microdomains (DRM) by self-aggregation during transport from the Golgi to the cell surface. The detergent insolubility of these aggregates stems from the properties of the lipids and their ability to form fluid vs. highly ordered membranes. DRM can be thought of as discrete lipid rafts in the general lipid milieu of the plasma membrane. These membrane patches, although rich in proteins, likely represent only 10-15% of the plasma membrane area. A good way to conceptualize these membrane microdomains is to visualize them as islands in the great lipid sea of the plasma membrane. At least two varieties of DRM exist on many cell surfaces (20) : One variety of cell surface DRM appear by electron microscopy (EM) as "flat" areas of the cell membrane and are referred to as G domains. The second variety of DRM are called caveolae and contain caveolin as well as other components described below (20) . By EM caveolae appear as striated 50-100 nm membrane "invaginations" coated with oligmerized caveolin. While many cell types have both caveolae and G domains, some cell varieties do not express caveolin (i.e. haemopoietic cells) and may only have G domains (21) . Caveolae Caveolae are distinctive DRM that appear as small non-coated plasmalemmal vesicles (Fig 2). These organelles are present in adipocytes, myocytes, endothelial cells, chondrocytes, osteoblasts as well as other cell types, and are formed by the oligmerization of the protein caveolin (a 22 kD phosphoprotein that is the major structural component of caveolae) in membranes with the lipid composition of DRM. As outlined below caveolae are implicated in molecular transport processes and signal transduction (22-26) . Caveolae in Molecular Transport: Caveolae are linked in a grape-like branching structure that penetrates deep within a cell, thus, conventional EM cannot be used to determine if caveolae are dynamic structures or remain attached to the cell membrane. Despite these problems, accumulating evidence makes it clear that caveolae are transport structures. This evi-dence includes the discovery that proteins involved in vesicular transport are found in caveolae (e.g. annexin, SNAP, NSF and VAMP-2) (26) . Caveolae in Signal Transduction: Many receptors and signal transduction molecules have been localized to caveolae these include: PDGF receptors (23-25) , GTP-binding proteins (26, 27) , Src family tyrosine kinases, PI3-kinase, PLC , PKC á and â (23) . Caveolin itself has been shown to associate with a variety of signal transduction molecules including GTP-binding proteins (27) , and Src family protein kinases (27) . It has been suggested that caveolin is a negative regulator of signaling molecule activity and may act as a scaffold' to which disparate signaling elements can attach in their inactive state (27) . Given the abundance of signaling molecules and receptors found in caveolae and DRM it has been suggested that they are pre-assembled signal transduction complexes. Caveolae and other DRM are loci for signal molecule communication because they are loci for signal molecule accumulation. To emphasize the role of caveolae in cell signaling we will use an instructive example of a growth factor receptor (platelet derived growth factor receptor: PDGFR) and explore the evidence that caveolae regulate its signal transduction. The PDGFR is a particularly interesting example as it is expressed on bone cells and bone cells are responsive to PDGF. PDGF receptors (PDGFR) in endothelial cells are associated with caveolae (23-25) , and require intact caveolae for maximum function (23) . In osteoblasts, PDGF treatment causes the phosphorylation of PDGFR in caveolin-enriched membranes and in the membrane at large (Solomon et al., 2000, submitted). It has also been shown that components of the PDGFR signaling cascade (e.g. Raf, Erk, Src) are present in caveolae of endothelial cells (23-25) , cardiomyocytes (28) , and osteoblasts (Solomon et al., 2000, submitted). PDGF treatment of endothelial cells leads to the activation of Erk in caveolae, and caveolae in isolation respond to PDGF by increasing tyrosine phosphorylation and activating Erk (25) . In osteoblasts, treatment of cells with PDGF causes translocation of Raf and Erk2 from the caveolin-enriched membranes to other areas of the cell (Solomon et al., 2000, submitted). Thus, caveolae are important regulators of PDGFR signaling in various cell types including osteoblasts. What Value is this Information in Understanding Osteoblast Function? How will the characterization of pre-assembled signaling complexes and caveolae provide fundamental new knowledge pertinent to the function of osteoblasts? How may this knowledge enable us to manipulate the function of osteoblasts for the development of new treatment protocols and improvement of current therapeutic approaches to bone disease? PDGF is an important growth factor for osteoblasts that regulates replication and gene expression, furthermore PDGF already is being used to stimulate bone repair in human clinical studies (29) . PDGFR is found in the pre-assembled signaling complexes of osteoblasts, endothelial cells, and myocytes, and receptor localization to these complexes is critical for function. Our recent observations regarding caveolae in osteoblasts reveal new targets for potentially enhancing the positive actions of PDGF while diminishing the negative ones. We may now address questions that previously we did not even know to ask: Are some of the multiple actions of PDGF on osteoblasts delivered by receptors regulated by their distribution into the plasma membrane at large vs. pre-assembled signaling complexes? Can we use this knowledge to promote desired clinical effects of PDGF while discouraging deleterious side-effects? Do the caveolae of osteoblasts regulate signaling from other growth factor receptors? Are the osteogenic effects of cholesterol-low-ering statin drugs (lovastatin, simvastatin) (30) explained by modulated signaling in cholesterol-rich caveolae? Answering these queries is part of our on-going and long-term research goals. Summary The discovery of new signaling pathways in osteoblasts and osteoclasts provide a window for understanding the complex interactions of these cells in bone. This new knowledge establishes an exciting foundation for future research into the pharmacological control of bone cells in health and disease. |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| HOJ HOME | Chiefs Reports | Osgood Day | Scientific Articles | Alumni Association |
| References | |
| 1. | Rodan GA, Martin TJ.Role of osteoblasts in hormonal control of bone resorption--a hypothesis. Calcif Tis Int 1981; 33:349-51. |
| 2. | Yasuda H, Shima N, Nakagawa N, et al.Osteoclast differentiation factor is a ligand for osteoprotegerin/osteoclastogenesis-inhibitory factor and is identical to TRANCE/RANKL. Proc Natl Acad Sci U S A 1998; 95:3597-602. |
| 3. | Anderson DM, Maraskovsky E, Billingsley WL, et al. A homologue of the TNF receptor and its ligand enhance T-cell growth and dendritic-cell function. Nature 1997; 390:175-9. |
| 4. | Wong BR, Josien R, Choi Y. TRANCE is a TNF family member that regulates dendritic cell and osteoclast function. J Leukoc Biol 1999; 65:715-24 |
| 5. | Simonet WS, Lacey DL, Dunstan CR, et al. Osteoprotegerin: a novel secreted protein involved in the regulation of bone density [see comments]. Cell 1997; 89:309-19. |
| 6. | Yasuda H, Shima N, Nakagawa N, et al. Identity of osteoclastogenesis inhibitory factor (OCIF) and osteoprotegerin (OPG): a mechanism by which OPG/OCIF inhibits osteoclastogenesis in vitro. Endocrinology 1998; 139:1329-37. |
| 7. | Lacey DL, Timms E, Tan HL, et al. Osteoprotegerin ligand is a cytokine that regulates osteoclast differentiation and activation. Cell 1998; 93:165-76 |
| 8. | Lee SK, Lorenzo JA. Parathyroid hormone stimulates TRANCE and inhibits osteoprotegerin messenger ribonucleic acid expression in murine bone marrow cultures: cor-relation with osteoclast-like cell formation. Endocrinology 1999; 140:3552-61. |
| 9. | Thomas RJ, Guise TA, Yin JJ, et al. Breast cancer cells interact with osteoblasts to support osteoclast formation. Endocrinology 1999; 140:4451-8. |
| 10. | Teitelbaum SL, Tondravi MM, Ross FP. Osteoclasts, macrophages, and the molecular mechanisms of bone resorption. J Leukoc Biol 1997; 61:381-8. |
| 11. | Hynes RO. Integrins: versatility, modulation, and signaling in cell adhesion. Cell 1992; 69:11-25. |
| 12. | Leu C-T, Wesolowski G, Nagy R, Francis B, Rodan GA, S.B.. R. Osteoclasts have over 107 high affinity echistatin binding sites (RGD-integrins). Vol. 12. Bone Min Res, 1997:S416. |
| 13. | Rodan SB, Rodan GA. Integrin function in osteoclasts. J Endocrinol 1997; 154 Suppl:S47-56. |
| 14. | Yamamoto M, Fisher JE, Gentile M, et al. The integrin ligand echistatin prevents bone loss in ovariectomized mice and rats. Endocrinology 1998; 139:1411-9. |
| 15. | Engleman VW, Nickols GA, Ross FP, et al. A peptidomimetic antagonist of the alpha(v)beta3 integrin inhibits bone resorption in vitro and prevents osteoporosis in vivo [see comments]. J Clin Invest 1997; 99:2284-92 |
| 16. | McHugh KP, Hodivala-Dilke K, Zheng MH, et al. Mice lacking beta3 integrins are osteosclerotic because of dysfunctional osteoclasts [In Process Citation]. J Clin Invest 2000; 105:433-40. |
| 17. | Chiba M, Teitelbaum SL, Cao X, Ross FP.Retinoic acid stimulates expression of the functional osteoclast integrin alpha v beta 3: transcriptional activation of the beta 3 but not the alpha v gene. J Cell Biochem 1996; 62:467-75. |
| 18. | Kitazawa S, Ross FP, McHugh K, Teitelbaum SL. Interleukin-4 induces expression of the integrin alpha v beta 3 via transactivation of the beta 3 gene. J Biol Chem 1995; 270:4115-20 |
| 19. | Inoue M, Namba N, Chappel J, Teitelbaum SL, Ross FP. Granulocyte macrophage-colony stimulating factor reciprocally regulates alphav-associated integrins on murine osteoclast precursors. Mol Endocrinol 1998; 12:1955-62. |
| 20. | Schnitzer JE, McIntosh DP, Dvorak AM, Liu J, Oh P. Separation of caveolae from associated microdomains of GPI-anchored proteins [see comments]. Science 1995; 269:1435-9. |
| 21. | Fra AM, Williamson E, Simons K, Parton RG. Detergent-insoluble glycolipid microdomains in lymphocytes in the absence of caveolae. J Biol Chem 1994; 269:30745-8. |
| 22. | Anderson RGW, Kamen BA, Rothberg KG, Lacey SW. Potocytosis: sequestration and transport of small molecules by caveolae. Science 1992; 255:410-411. |
| 23. | Liu J, Oh P, Horner T, Rogers RA, Schnitzer JE. Organized endothelial cell surface signal transduction in caveolae distinct from glycosylphosphatidylinositol-anchored protein microdomains. J Biol Chem 1997; 272:7211-22. |
| 24. | Liu P, Ying Y, Ko YG, Anderson RG. Localization of platelet-derived growth factor-stimulated phosphorylation cascade to caveolae. J Biol Chem 1996; 271:10299-303. |
| 25. | Liu P, Ying Y, Anderson RG. Platelet-derived growth factor activates mitogen-activated protein kinase in isolated caveolae. Proc Natl Acad Sci U S A 1997; 94:13666-70. |
| 26. | Schnitzer JE, Liu J, Oh P. Endothelial caveolae have the molecular transport machinery for vesicle budding, docking, and fusion including VAMP, NSF, SNAP, annexins, and GTPases. J Biol Chem 1995; 270:14399-404 |
| 27. | Li S, Couet J, Lisanti MP. MP. Src tyrosine kinases, Galpha subunits, and H-Ras share a common membrane-anchored scaffolding protein, caveolin. Caveolin binding nega-tively regulates the auto-activation of Src tyrosine kinases. J Biol Chem 1996; 271:29182-90. |
| 28. | Rybin VO, Xu X, Steinberg SF. Activated protein kinase C isoforms target to cardiomyocyte caveolae : stimulation of local protein phosphorylation. Circ Res 1999; 84:980-8. |
| 29. | Howell TH, Fiorellini JP, Paquette DW, Offenbacher S, Giannobile WV, Lynch SE. A phase I/II clinical trial to evaluate a combination of recombinant human platelet-derived growth factor-BB and recombinant human insulin- like growth factor-I in patients with periodontal disease. J Periodontol 1997; 68:1186-93. |
| 30. | Mundy G, Garrett R, Harris S, et al. Stimulation of bone formation in vitro and in rodents by statins [see comments]. Science 1999; 286:1946-9. |
|
TOP OF PAGE | HOJ HOME |
| HOJ HOME | Chiefs Reports | Osgood Day | Scientific Articles | Alumni Association |