T-cell Co-stimulators in Osteolysis Following Total Hip Replacement
Mahito Kawashima, PhD, K Hayata, PhD, Harry E. Rubash, MD, and Arun S. Shanbhag, PhD
Biomaterials Lab, Massachusetts General Hospital
Introduction
 Interfacial tissues surrounding failed joint replacements
have provided crucial insight into the mechanisms of periimplant
bone loss and implant loosening. Using high throughput
protein chips, we recently reported high levels of T-cell chemokines:
interferon-?-inducible protein of 10KDa (IP-10) and
monokine induced by interferon-? (MIG) in osteolytic tissues
(1). The question posed is: if activated T-cells are recruited to
the inflammatory site, are other co-stimulators and modulators
of antigen presentation and immunological synapse formation
(B7-1, B7-2 and CD-28) also upregulated? We tested for the
gene expression of these modulators, as well as IP-10, MIG and
cytotoxic T-lymphocyte antigen 4 (CTLA-4) using RT-PCR .
Interfacial tissues surrounding failed joint replacements
have provided crucial insight into the mechanisms of periimplant
bone loss and implant loosening. Using high throughput
protein chips, we recently reported high levels of T-cell chemokines:
interferon-?-inducible protein of 10KDa (IP-10) and
monokine induced by interferon-? (MIG) in osteolytic tissues
(1). The question posed is: if activated T-cells are recruited to
the inflammatory site, are other co-stimulators and modulators
of antigen presentation and immunological synapse formation
(B7-1, B7-2 and CD-28) also upregulated? We tested for the
gene expression of these modulators, as well as IP-10, MIG and
cytotoxic T-lymphocyte antigen 4 (CTLA-4) using RT-PCR .
MATERIALS AND METHODS
Clinical
Interfacial tissues were harvested from patients (n=14;
mean age = 66y) undergoing revision surgery for aseptic loosening
of their femoral total hip replacement (THR). Capsular
tissues from patients with end-stage osteoarthritis (OA) undergoing
primary THR (n=14, mean age = 67y) provided control
comparisons. In the operating room, tissues were flash frozen
in liquid nitrogen and subsequently homogenized and had their
mRNA extracted.
RNA Extraction and RT-PCR
Total RNA was extracted from 1cm3 of periprosthetic tissue
by homogenization in 1.6mL TRIzol reagent (Invitrogen,
Paisley, UK). RNA was extracted using established protocol and
cleaned using RNeasy® Mini kit (Qiagen, Valencia, CA). RNA
samples were stored at (-)76 oC.
PCR amplification was performed in 50 µl reactions containing
20 µl Eppendorf® Master Mix (2.5x) (Eppendorf AG,
Hamburg, Germany), 2 µl cDNA and each oligonucleotide
primer at 0·5 µM. Oligonucleotide primers were designed
according to previously published sequences for IP-10 (2),
MIG (3), B7-1/CD80 and B7-2/CD86 (4), CD28 (5), and CTLA-
4/CD152 (6). GAPDH was used as a housekeeping gene to test
integrity of sample cDNA. Data was analyzed using both the F
statistic and Student’s t-test. A p-value of <0.05 was considered significant.
RESULTS
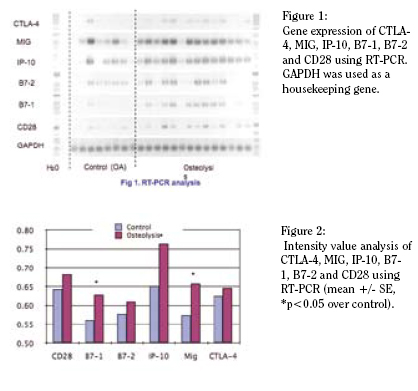
Compiled PCR results are shown in Figure 1 and the intensity
values are plotted in Figure 2. In most cases of implant
loosening significant levels of MIG and IP-10 were detected.
This finding concurs with our protein studies indicating significant
release of these two important T-cell chemokines (1). We
also detected strong expression of T-cell modulators: B7-1, B7-
2, CD28 in osteolysis samples and to a lesser extent in OA cases.
Interestingly, CTLA-4, a repressor of the T-cell activation, was
also upregulated in a few osteolysis cases (not significant).
DISCUSSION
IP-10 and MIG are produced by IFN-? stimulated monocytes,
macrophages and antigen experienced T cells and are a
chemoattractant for activated Th1 cells and natural killer (NK)
cells. They also share common receptor. In this work we have
demonstrated that these higher protein levels are associated
with a concomitant upregulation of gene expression. These
findings suggest that antigen is being presented and activated
T-cells are indeed recruited to the inflammatory site in osteolysis.
Additional co-stimulators such as B7-1, B7-2 and CD28
are also upregulated – these are required for the development
of the immunological synapse between the antigen-presenting
cell (APC) and the T-cell. B7-1 and B7-2 have largely overlapping
functions. They provide important co-stimulatory signals
to augment and sustain a T cell response via an interaction
with CD28. B7-2 is expressed constitutively at low levels on
APCs and is rapidly upregulated in immune responses, whereas
B7-1 is inducibly expressed later after activation. In the present
study, we demonstrated a significantly higher level of B7-1 gene
expression in osteolytic tissues, compared with B7-2 expression.
These results suggest that osteolysis may represent a late
stage in the immune response.
Because important T-cell mediators such as IL-2, INF-?,
IL-1, and TNF-a are not detected in protein arrays, the immune
response appears to have been aborted (1). Notably, mRNA
expression of CTLA-4 is upregulated in some cases of osteolysis.
CTLA-4 is an integrin expressed on activated T-cells and
inhibits IL-2 secretion from T-cells (7). This is additionally supported
by high protein levels of soluble intercellular adhesion
molecule-1 (sICAM-1) which serves as a soluble receptor for
the leukocyte integrin, lymphocyte function-associated antigen
(LFA-1)/CD18. While LFA-1 facilitates leukocyte adhesion and
migration across the endothelium, sICAM-1 binding to LFA
disrupts the immune synapse and prevents propagation of a
robust T-cell mediated immune response (8).
SUMMARY
T-cell co-stimulators, CD28, B7-1 and B7-2 are crucial for
facilitating and stabilizing the immunological synapse, and are
expressed in osteolytic tissues. While the underlying mechanisms
are still being clarified, these results indicate that the
participation of an immunological reaction, alongside the well
described macrophage-mediated response to foreign body, is
important in leading to osteolysis and aseptic loosening of total
joint replacements.
Acknowledgements: MGH Orthopaedics for financial support
Mahito Kawashima Ph.D. is a member of the Biomaterials Lab at Massachusetts General Hospital.
K Hayata Ph.D. is a member of the Biomaterials Lab at Massachusetts General Hospital.
Harry E. Rubash M.D. is Chief of the Orthopedic Department at Massachusetts General Hospital.
Arun S. Shanbhag Ph.D., MBA is Director of the Biomaterials Lab at Massachusetts General Hospital and Assistant Professor of Orthopedic Surgery at Harvard Medical School.
Address correspondence to:
Arun Shanbhag, Ph.D, MBA
GRJ 1115, 55 Fruit St.
Boston, MA 02114
References:
- Shanbhag AS, Kaufman AM, Agarwal S, et al.. Critical Insights into osteolysis using protein microarrays: The importance of IL-6 and T-cell Activation. Trans Orthop Res Soc 2005; 30:148.
- Gasper NA, Petty CC, Schrum LW, et al.. Bacterium-induced CXCL10 secretion by osteoblasts can be mediated in part through toll-like receptor 4. Infect Immun. 2002 Aug;70(8):4075-82.
- Raju,R, Malloy,A, Shah,T, et al.. Alloimmune induction of endothelial cell-derived interferon-gamma-inducible chemokines: Transplantation. 75:1072-4 (2003)
- Kiefer R, Dangond F, Mueller M, et al.. Enhanced B7 costimulatory molecule expression in inflammatory human sural nerve biopsies: J Neurol Neurosurg Psychiatry. 69: 362-368 (2000)
- Xu KL, Zhang Y, Pan XY, Lu QX. Inhibiting the expression of CD28 costimulatory molecule on human lymphocytes by special siRNA: Clin Med J. 118: 480-486 (2005)
- Murata K, Dalakas MC. Expression of the costimulatory molecule BB-1 the ligands CTLA-4 and CD28 and their mRNA in inflammatory myopathies: Am J Pathol. 155:453-60 (1999)
- Walunas TL, Lenschow DJ, Bakker CY, et al.. CTLA-4 can function as a negative regulator of T cell activation: Immunity. 1: 405-413 (1994).
- Rieckmann P, Michel U, Albrecht M, et al.. Soluble forms of intercellular adhesion molecule-1 (ICAM-1) block lymphocyte attachment to cerebral endothelial cells: Neuroimmunol. 60: 9-15 (1995).
|