
Original Submissions
Current Issue: Volume 14 |  PDF
PDF
Advances in Single-cell Tracking of Mesenchymal Stem Cells (MSCs) During Musculoskeletal Regeneration
1Children’s Hospital Boston, Harvard School of Dental Medicine, Boston, MA
2Department of Medicine, Center for Regenerative Therapeutics & Department of Medicine, Division of Biomedical Engineering, Brigham and Women’s Hospital, Harvard Medical School, Harvard Stem Cell Institute, Harvard-MIT Division of Health Sciences and Technology, Massachusetts Institute of Technology, Cambridge, MA
3Advanced Microscopy Group, Wellman Center for Photomedicine and Center for Systems Biology, Massachusetts General Hospital, Harvard Medical School, Boston, MA
PDF
Abstract
The combined cost of treatment and lost wages due to musculoskeletal disease approaches one trillion dollars a year. It is crucial to develop new treatments to alleviate the cost and morbidity of these diseases. Due to their multilineage differentiation potential and potent trophic activities, mesenchymal stem cells (MSCs) can promote multiple types of connective tissue regeneration, immune suppression, down-regulation of inflammation, and neovascularization. MSCs can supply many cell types needed for musculoskeletal tissue regeneration and modulate inflammation. Therapeutically, endogenous MSCs can respond locally or home to injuries from distant sites. Culture expanded MSCs can be infused systemically or injected locally. Understanding how MSCs respond to connective tissue injury in these different situations is key to developing regenerative therapeutics. Current cell-tracking methods, like bioluminescence, do not have enough spatial-temporal resolution to determine how MSCs contribute to regeneration. To visualize MSCs during tissue regeneration, we applied fluorescence confocal and multi-photon in vivo imaging to track single-cells in various mouse models of musculoskeletal disease. These models can be used to elucidate factors that influence MSC recruitment, migration, and proliferation during musculoskeletal regeneration and lead to the creation of novel regenerative therapies.
Introduction
Musculoskeletal diseases are the most commonly reported health conditions in the United States.1 These diseases include various forms of arthritis, congenital deformities and anomalies, fractures, and pain associated with the back, neck, or intervertebral disks.1 Millions of surgeries are performed every year to correct musculoskeletal diseases, yet a high percentage fail to obtain a satisfactory outcome.2-5 For example, only 64% of patients treated surgically for lumbar spinal stenosis reported good-to-excellent outcomes.4 Treatments and lost wages due to musculoskeletal diseases are a significant burden on society, representing 7.7% gross domestic product (~$849,000,000,000) for 2002 to 2004.1 Therefore, new therapies that reduce the cost and morbidity of musculoskeletal diseases are in great demand.
MSCs have been the focus of widespread attention in recent years, and are being studied in over one hundred clinical trials world-wide.6 In addition to their obvious potential for treatment of musculoskeletal diseases, MSCs have demonstrated promise as a cell therapy in many pre-clinical and early-stage clinical trials for a range of diseases, including diabetes, cardiac disease, bone marrow transplant-associated GVHD, and osteogenesis imperfecta.6,7 Although proven to be safe, efficacy in late stage clinical trials has not met expectations.8,9 This has led some to develop strategies that can enhance the potency of MSCs before infusion, including activation or transfection of cells before infusion, and cell surface engineering.10,11 A critical challenge in the development of these enhancement strategies is quantification of in vivo MSC homing and therapeutic efficiency. To address this challenge, we applied in vivo confocal and multi-photon microscopy, a powerful singe-cell detection and evaluation technique.
MSC use in pre-clinical and clinical models began with their discovery in the 1960’s during bone marrow transplant experiments that led to the hypothesis that a cell type existed within the bone marrow that could differentiate into osteoblasts, aid in the development of sinusoidal structures, and support hematopoiesis in ectopic sites.12 These skeletal stem cells were first purified from bone marrow based on adherence to tissue culture plastic and were re-named MSCs for their ability to differentiate into adipocytes, chondrocytes, and osteoblasts.13,14 MSCs have been well characterized in vitro, and are defined as being tissue culture plastic adherent; positive for CD105, CD73, and CD90 antigens; negative for HLA-DR, and CD45, CD34, CD14, CD19 antigens; and able to differentiate into osteoblasts, adipocytes, and chondrocytes.15 Recently, it has been suggested that MSCs are perivascular cells in vivo. This has raised many interesting questions about how pericytes and adventitial cells respond to acute bone injury and if perivascular cells in non-bone tissues are also MSCs.16-18
Our research describes the ability of in vivo confocal and multi-photon microscopy to quantify the behavior of exogenous engineered MSCs and evaluate endogenous perivascular cell response to bone injury. Our results contribute to our understanding of how exogenous MSCs interact with diseased tissue and how endogenous perivascular cells respond to musculoskeletal injury. These results may help improve current cell therapies and lead to the development of novel therapeutics.
Results
Using the computer model, the two evaluative methods, MADC and MADG, produced greatly varying results. MADG significantly exceeded MADC values, which stresses the severity of the malalignment. Higher angles of hindfoot deformity were associated with greater MADG from anatomical axis. In future studies using the cadaveric model, we expect to find that load measured at the ground reaction point is closer to the actual force applied when compared with load measured at the center of the ankle. This difference between measured load transmission at the center of the ankle and the ground reaction point should be greater for each specimen after hindfoot deformity has been simulated. Based on our computer model findings, we also expect that knees implanted using MADC to plan realignment will demonstrate more uneven loading at the medial and lateral knee joint when compared to those implanted using MADG for surgical planning.
Materials and Methods
Video-rate Laser Scanning Confocal And Multi-photon Hybrid Imaging System
Our video-rate laser scanning confocal and multi-photon hybrid imaging system is designed specifically for live animal cell-tracking and molecular imaging studies (Figure 1).19 The system is equipped with three monochrome lasers and a Ti:Sapphire laser. The Ti:Sapphire laser can be tuned between 710-920 nm and can be used for two-photon microscopy. Enhanced Green Fluorescent Protein (EGFP) can be imaged by either confocal (λex = 491 nm, 509-547 nm detection) or two-photon microscopy (λex = 920, 505-575 nm detection). The system is designed to image EGFP and three other standard fluorescent channels: DsRed (λex = 532 nm, 573-613 detection), DiD (λex =633 nm, 667-722 nm detection), and DiR (λex = 750 nm, >770 nm detection); as well as collagen/bone using Second Harmonic Generation (SHG, λinc = 880 nm, λscat = 440 nm). The Ti:Sapphire laser is used to excite both SHG and 2-photon imaging modalities since these are multi-photon processes that require high power and short pulse duration, ~100 fs. A rotating polygon mirror in series with a galvanometer mirror allows the x-y laser scan to be performed at video-rate (30 frames/sec).
Several aspects of this system allow us to track single-cells in real-time within inflamed tissue and calvarial bone marrow inside live animals. First, up to three confocal fluorescent channels or two confocal fluorescent and one two-photon channel can be recorded simultaneously. This allows us to image cells, blood vessels, and bone simultaneously (Figure 2A). Also, cells expressing specific markers can be visualized by infusing fluorescent antibodies intravenously or subcutaneously.20 Second, we can probe skin to a depth of ~250 µm and calvarial bone marrow to a depth of ~150 µm. The spatial resolution is sufficient to determine distances between cells, bone surface, and blood vessels. This is particularly useful when defining the stem cell niche and observing extravasation of cells infused in the blood stream (Figure 2B).10,21-23 Third, by utilizing the video-rate scanning capabilities, heartbeat and breathing artifacts are minimized, and it is possible to visualize circulating MSCs interacting with endothelium (Figure 2C). From these video-rate images, the rolling velocities of MSCs can be quantified.11
MSC Homing
MSCs have the ability to home to bone marrow and sites of inflammation/injury. To assess homing, we used two methods of in vivo imaging, i.e. bone marrow and inflamed ear imaging. Studies were performed in accordance with US NIH guidelines for care and use of animals under approval of the Institutional Animal Care and Use Committees of Massachusetts General Hospital, Children’s Hospital Boston, and Harvard Medical School. Intravital imaging of bone marrow and inflamed ears was performed as previously described.10,24 For delineation of vasculature during imaging, fluorescent-conjugated dextran (2 x 106 Da; Invitrogen) was infused intravenously just prior to imaging. For bone marrow MSC homing studies, 106 DiD (Invitrogen) labeled MSCs were infused intravenously twenty-four hours prior to imaging. For inflamed ear MSC homing studies, inflammation was induced via injection of E. coli lipopolysaccharide (LPS) into the ear pinna of the right ear, while the left ear received saline as a control. Twenty-four hours after induction of inflammation, 106 MSCs were infused intravenously. Imaging was then performed 24 hours after MSC infusion.
Perivascular Cell Proliferation And Migration After Osseous Injury
To reliably image perivascular cells in vivo requires markers that have high specificity to pericytes and adventitial cells in the tissue of interest. CD146 antigen, nestin, and alpha-smooth muscle actin (SMA) are the most specific candidates characterized in bone marrow.21,26,27 To observe pericyte and adventitial cell proliferation, migration, and homing during osseous regeneration, we imaged calvarial defects created in transgenic mice expressing EGFP under the SMA promoter. In these mice, SMA-EGFP cells are found in perivascular locations within mouse calvarial bone marrow. Figure 4A shows a large area map of the calvarial bone in which perivascular cells are associated with sinusoidal microvessels within the parasagittal bone marrow regions. In this mouse model, vascular smooth muscle cells can be visualized wrapped around arterioles (Figure 4A, arrowheads). Non-critical osseous defects were created in parasagittal bone marrow containing regions using a dental drill (Figure 4A, white boxes) and characterized using SHG imaging (Figure 4B). Briefly, the scalp was surgically retracted and calvarial defects were created using a dental drill. Between daily time-lapse imaging sessions, the scalp was sutured and mice were allowed to fully recover from anesthesia.
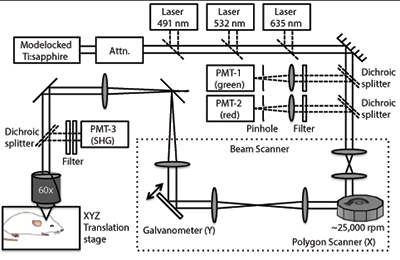 Schematic of in vivo confocal fluorescence and multi-photon microscope. A rotating 36-faceted polygonal mirror creates the X-scan and a galvanometer-mounted mirror creates the Y-scan. The mode locked Ti:sapphire laser provides wavelength-tunable multi-photon illumination. The anesthetized animal is placed under the objective in a heated 3D-translation stage, which has micron resolution.
Schematic of in vivo confocal fluorescence and multi-photon microscope. A rotating 36-faceted polygonal mirror creates the X-scan and a galvanometer-mounted mirror creates the Y-scan. The mode locked Ti:sapphire laser provides wavelength-tunable multi-photon illumination. The anesthetized animal is placed under the objective in a heated 3D-translation stage, which has micron resolution.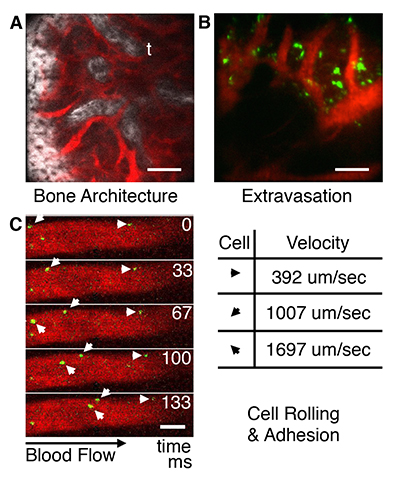 Basic capabilities of in vivo microscope. (A) Calvarial bone microstructures (white) and blood vessels (red); t = trabecula. (B) Visualization of MSC (green) extravasation in bone marrow. (C) Images of MSCs flowing through blood vessels acquired at 30 frames/second can be used to quantify cell velocity for cell rolling and adhesion studies. Bar = 100 μm.
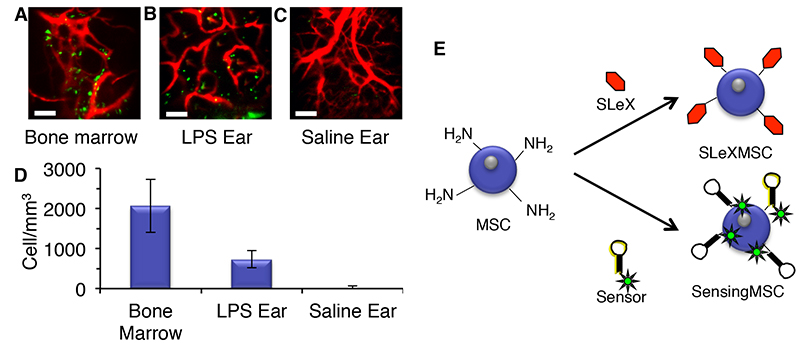
Basic capabilities of in vivo microscope. (A) Calvarial bone microstructures (white) and blood vessels (red); t = trabecula. (B) Visualization of MSC (green) extravasation in bone marrow. (C) Images of MSCs flowing through blood vessels acquired at 30 frames/second can be used to quantify cell velocity for cell rolling and adhesion studies. Bar = 100 μm.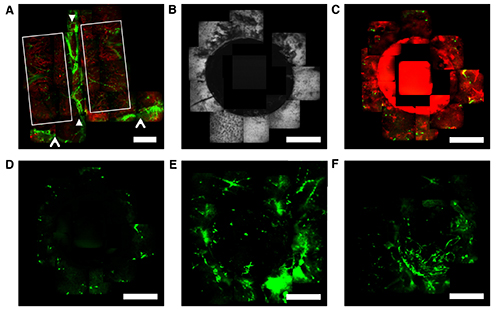 MSC homing to bone marrow and inflamed tissue twenty-four hours after systemic infusion. (A) MSC homing to calvarial bone marrow. (B) MSC homing to LPS treated ear tissue. (C) Lack of MSC in saline treated ear tissue. MSC (green); Blood vessels (red); Bar = 100 μm. (D) Cell density per cubic mm. (E) Chemical engineering of MSCs with homing ligands and molecular sensors.
MSC homing to bone marrow and inflamed tissue twenty-four hours after systemic infusion. (A) MSC homing to calvarial bone marrow. (B) MSC homing to LPS treated ear tissue. (C) Lack of MSC in saline treated ear tissue. MSC (green); Blood vessels (red); Bar = 100 μm. (D) Cell density per cubic mm. (E) Chemical engineering of MSCs with homing ligands and molecular sensors.All Rights Reserved. Permission For Use Required.
References
Results And Discussion
MSC Homing
As shown in Figure 3, high numbers of MSCs can be observed in bone marrow and inflamed ears (Figures 3A & 3B), but not in saline treated ears (Figure 3C). Images from these experiments were evaluated using ImageJ software (NIH). Thousands of MSCs per mm3 were found in calvarial bone marrow and inflamed ear tissue, while only ~22 MSCs per mm3 was observed in saline treated ears (Figure 3D).
After homing to damaged tissues, MSCs may secrete trophic factors or supply cell types that are necessary for tissue regeneration.25 Increasing the numbers of MSCs that home to injuries could potentially increase the efficacy of MSC therapies.10 We have developed methods to coat the surface of MSCs with homing ligands typically expressed on leukocytes, e.g. Sialyl Lewis X or SLeX. Our imaging methods demonstrated that engineered MSCs exhibit increased homing to inflamed tissue and slower rolling velocities on inflamed endothelium and (Figure 3E).11 We have also developed molecular sensors that can be placed on cell surfaces to sense the local microenvironment of MSCs that have homed to bone marrow (Figure 3E).23 We believe that these chemical engineering methods will enhance the efficacy of MSC therapies and could lead to discoveries about the make-up of the microenvironment that MSCs encounter in vivo.
Perivascular Cell Proliferation And Migration After Osseous Injury
Little is known about the process of osseous regeneration at the single-cell level. In general, repair and regeneration of osseous injuries depends on the recruitment, activation, and differentiation of competent adult MSCs as well as angiogenesis and production of reparative bone matrix. Osseous regeneration requires the spatial and temporal regulation of multiple cell types at various stages of differentiation. MSCs may be recruited from the local milieu, or mobilized to the injury from distant sites. These cells gradually advance into the core of the defect leaving a trail of new tissue. Our long-term goal is to generate a clear understanding at the single-cell level of osseous regeneration. This knowledge can be used to develop the potency of therapies for regenerative medicine and tissue engineering. Although the in situ identity of MSCs has been controversial, recent evidence suggests that perivascular cells, namely pericytes and adventitial cells, are the in vivo origin of MSCs.17,18,26 Therefore, we applied in vivo imaging techniques to visualize the response of perivascular cells during osseous regeneration.
Immediately after injury, a hematoma formed within the defect (Figure 4C). Time-lapse microscopy revealed a significant increase in EGFP signal and number of SMA-EGFP+ cells in bone marrow adjacent to the defect at two days post injury (Figure 4D & 4E). Three days post injury, the number of SMA-EGFP+ cells inside the defect increased. This data suggests that SMA-EGFP+ cells respond to osseous injury by proliferating and migrating into the defect.
SMA-EGFP+ cell dynamics during osseous regeneration. (A) Images of uninjured animal showing parasagittal regions outlined with white box, central vein and sagittal suture running top-to-bottom between closed arrowheads, and coronal vein and suture running left-to-right highlighted with open arrowheads. Vascular smooth muscle cells can be seen wrapped around arterioles associated with the central and sagittal sutures. Perivascular SMA-EGFP+ cells can be visualized in the parasagittal regions. (B) Representative images of calvarial bone surface after creation of 1.2 mm osseous defect. (C) Representative images of blood and SMA-EGFP signals after creation of osseous defect. (D-F) Time-lapse imaging of SMA-EGFP signal immediately, three days, and five days after osseous injury. SMA-EGFP (green); blood (red); bone (white); Bar = 500 μm.
All Rights Reserved. Permission For Use Required.Conclusion
We applied live-animal single-cell in vivo imaging techniques to characterize various aspects of MSC therapies and MSC-host interactions during inflammation and osseous regeneration. These imaging techniques have allowed us to demonstrate the improved homing of engineered MSCs, the feasibility of sensing the MSC microenvironment, and the endogenous perivascular SMA-EGFP response to osseous injury. This imaging technology will be helpful to understand the molecular mechanisms that MSCs use to tether and adhere to inflamed endothelium and transmigrate across endothelium. By characterizing the activities of MSCs during osseous regeneration, the mechanisms or specific cues that mediate the migration and differentiation may potentially be identified. These techniques could also be used to determine how bone-engineering strategies regulate MSC migration and differentiation.
Acknowledgements
This work was supported by the NIH grant# HL095722 to JMK and CDMRP-DOD grant# OR090192 to PVH. Primary human MSCs were obtained from the Center for Gene Therapy at Tulane University (grant P40RR017447, from National Center for Research Resources of the National Institute of Health).
References
